Coccolithophores and Calcareous Nannofossils
A Beginner’s Guide
Coccolithophores are among the most abundant and important microscopic organisms in the world's oceans. These single-celled algae produce complex calcium carbonate plates known as coccoliths, which surround the cell and form a coccosphere. Although invisible to the naked eye, coccolithophores play a fundamental role in marine ecosystems, the global carbon cycle, and the formation of marine sediments that preserve an extraordinary fossil record spanning over 200 million years.
This beginner’s guide provides a simple introduction to coccolithophores, calcareous nannoplankton, and calcareous nannofossils. It explains the differences between these commonly confused terms and introduces the structure of the coccolithophore cell, its life cycle, biomineralisation processes, and its role in the biological and carbonate carbon cycles. The aim is to provide students, newcomers to nannofossil studies, and interested readers with an accessible overview of these remarkable organisms and their significance in both modern oceans and the geological record.
The terms coccolithophores, calcareous nannoplankton, and calcareous nannofossils are often used interchangeably, but they refer to different biological and geological concepts. All of these belong to the kingdom Protista and the phylum Haptophyta, playing significant roles in global carbon fixation and in regulating Earth's climate.
Coccolithophores (living naked or coccosphere-bearing organisms) are unicellular, flagellated marine algae that inhabit the photic zone, which is the sunlit upper layer of the ocean. They can exist either as naked cells or as cells enclosed within a coccosphere made up of calcium carbonate (CaCO₃) plates known as coccoliths. Coccolithophores belong to the class Prymnesiophyceae.
Calcareous nannoplankton (the calcifying component of the nannoplankton) refers specifically to the living forms of coccolithophores that bear coccoliths, and they are classified within the classes Coccolithophyceae or Prymnesiophyceae.
Calcareous nannofossils consist of fossilised coccospheres and individual coccoliths produced by calcareous nannoplankton (coccolith-bearing coccolithophores), together with associated calcareous microfossils of similar size, such as discoasters and schizosphaerellids (nannoliths). The group also includes unrelated forms, such as pithonellids and didemnid ascidian spicules, which are traditionally studied alongside calcareous nannofossils because of their comparable size and occurrence in marine sediments.
The informal term nannoplankton (derived from the Greek prefixes nano- and nanno-, both meaning “dwarf”) was introduced by Lohmann (1908) to describe plankton ranging from 5 to 50 μm in size. Modern usage generally defines nannoplankton as plankton ranging from approximately 2 to 63 μm, encompassing organisms small enough to pass through the finest plankton nets. This size category includes a wide variety of marine microorganisms, such as coccolithophores, small diatoms, silicoflagellates, dinoflagellates, chrysophytes, cryptophytes, and other microscopic algae and protists. While the prefix nano- is now widely used in technology and physics, nanno- remains the preferred form in biological and palaeontological terminology. Among these organisms, coccolithophores are of particular importance because of their abundance, their role in marine carbon cycling, and their extensive fossil record.
Summary of differences:
Coccolithophores = living naked or coccosphere-bearing organisms
Calcareous nannoplankton = living coccosphere-bearing coccolithophores
Calcareous nannofossils = fossil remains of calcareous nannoplankton and associated calcareous fossils, regardless of biological affinity.
Coccolithophore cell
Although invisible to the naked eye, the coccolithophore cell is an extremely organized and complex living entity. Within a confined space of just a few micrometers in diameter, it contains the organelles essential for photosynthesis, respiration, growth, and reproduction. In many species, the cell is protected by a delicate covering made of calcium carbonate plates called coccoliths, forming a spherical sheath known as the cocososphere. Through the combined activities of billions of these cells, coccolithophores influence marine ecosystems, the global carbon cycle, and Earth's climate.
Most motile coccolithophores possess two flagella, which are thin, whip-like extensions that propel the cell through seawater. While the ecological significance of this motility is not fully understood, it is thought to help cells respond to environmental changes and maintain favorable conditions for growth and photosynthesis.
The two flagella are accompanied by a third appendage, the haptonema. Unlike the flagella, the haptonema does not appear to play a major role in propulsion. Although its functions are not fully understood, observations suggest that it is involved in sensing the surrounding environment, interacting with particles, and responding to external stimuli. In some haptophytes, the haptonema can coil rapidly, possibly as a defensive or sensory response.
Nucleus. The nucleus is the control centre of the coccolithophore cell. It contains the cell's genetic material (DNA), organised into chromosomes that carry the instructions for growth, photosynthesis, calcification, reproduction, and other cellular activities. By regulating gene expression, the nucleus coordinates the production of proteins required for coccolith formation and cellular metabolism. During cell division, DNA is replicated and passed to daughter cells, ensuring genetic continuity. Over evolutionary timescales, mutations in nuclear DNA have contributed to the remarkable diversity of coccolith morphology observed in both living species and the fossil record.
Mitochondrion. The mitochondrion is responsible for producing most of the cell's usable energy in the form of adenosine triphosphate (ATP). Through cellular respiration, it converts energy stored in organic compounds into a form that can be used to power cellular processes. In coccolithophores, this energy supports growth, maintenance, cell division, motility in flagellated stages, and the energy-demanding process of coccolith production. Mitochondria also participate in carbon metabolism and help regulate the intracellular conditions required for normal cellular function and biomineralisation.
Chloroplasts. Chloroplasts are the sites of photosynthesis, the process by which coccolithophores use sunlight to convert carbon dioxide (CO₂) and water into organic compounds that support growth and reproduction. Most coccolithophores possess two chloroplasts, each containing chlorophyll pigments, thylakoid membranes, and a pyrenoid. Through photosynthesis, chloroplasts provide the organic carbon required for cellular metabolism and contribute significantly to marine primary productivity and the global carbon cycle. The uptake of CO₂ by chloroplasts is also closely linked to the processes of calcification and carbon export in the oceans.
Plasma Membrane. The plasma membrane forms the interface between the coccolithophore cell and the surrounding seawater. It regulates the exchange of nutrients, carbon dioxide, calcium ions, and other dissolved substances required for photosynthesis, growth, and calcification. By controlling the transport of these materials, the plasma membrane helps maintain the internal conditions necessary for the formation of coccoliths and the normal functioning of the cell.
Golgi Apparatus. The Golgi apparatus plays a central role in coccolith production. It packages and transports the organic molecules required for coccolith formation and gives rise to the coccolith vesicles in which biomineralisation occurs.
Organic Scales. Organic scales form a thin layer between the periplast and the coccosphere. They provide a template upon which coccoliths are positioned and may play an important role in determining coccolith spacing, orientation, and the overall dimensions of the coccosphere.
Biomineralisation. During biomineralisation, coccoliths are formed within specialised coccolith vesicles, where an organic matrix precisely controls crystal nucleation, growth, and morphology. After their formation, the coccoliths are transported to the cell surface and incorporated into the coccosphere. The size and arrangement of the underlying organic scales may influence the spacing and organisation of coccoliths and, consequently, the overall dimensions and architecture of the coccosphere.
Coccosphere. The coccosphere is a covering of coccoliths that surrounds the coccolithophore cell. It is thought to provide protection against grazers, viruses, and mechanical damage while also shielding the cell from excessive light and ultraviolet radiation. The coccosphere may influence buoyancy, regulate interactions between the cell and the surrounding seawater, and contribute to the transport of carbon to the deep ocean after the cell dies. Although its precise functions remain incompletely understood, the coccosphere is widely regarded as an adaptation that enhances survival in the marine environment.
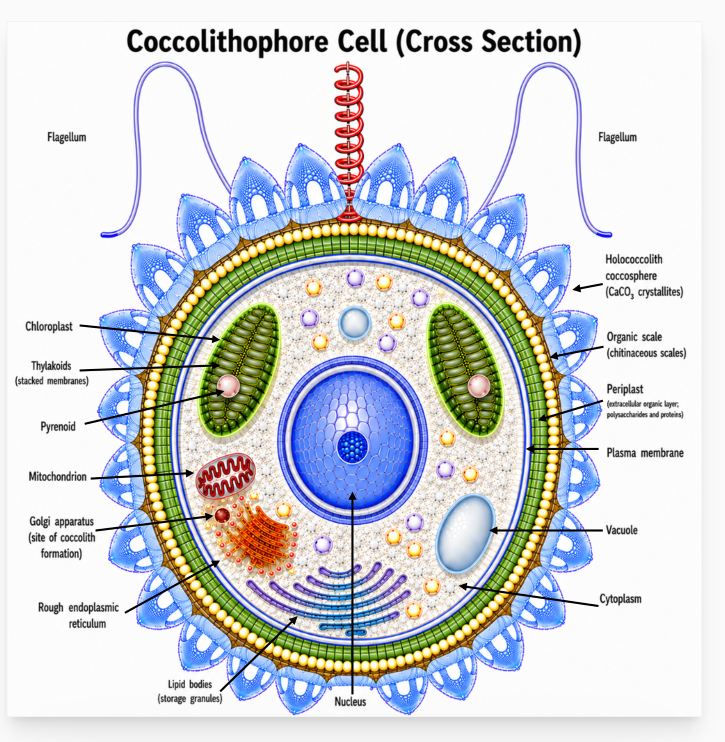

Schematic cross-section of a holococcolithophore cell. The reconstruction of the coccosphere is inspired by published photographs of Calyptrosphaera heimdaliae in Young et al. (2003, Plate 45.3) and by specimens from the research collection of Annelies Kleijne as documented in Nannotax. The internal cell structures are schematic and are intended to illustrate the principal organelles and extracellular layers of a coccolithophore cell.
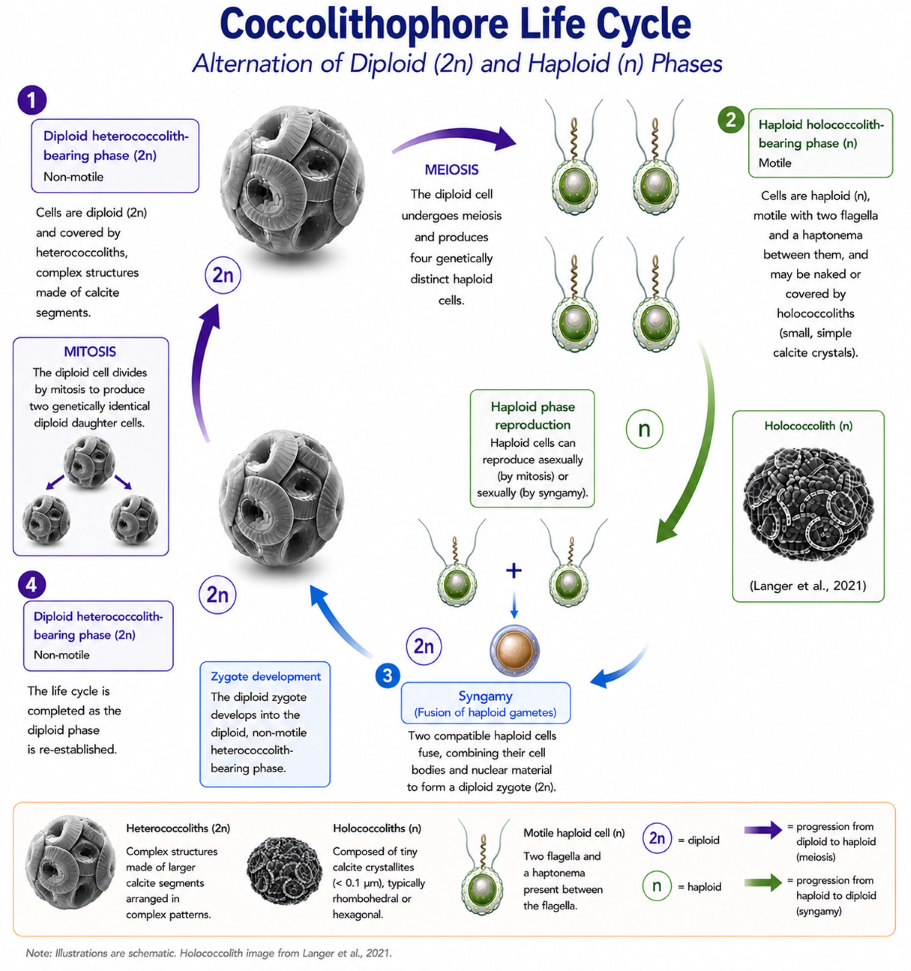
Coccolithophore Life Cycle
Coccolithophores undergo a life cycle that alternates between diploid and haploid phases, which is commonly referred to as a dimorphic life cycle. Parke and Adams (1960) were the first to propose that holococcoliths and heterococcoliths represent different stages in the life cycle of the same species. This interpretation was later supported and expanded by subsequent studies, including those by Rowson et al. (1986). This discovery significantly enhanced our understanding of coccolithophore biology by demonstrating that forms previously considered to be separate species might actually represent distinct life-cycle stages of a single organism.
Coccolithophores reproduce asexually through mitosis, where one diploid cell divides to create two genetically identical diploid daughter cells. They can also undergo meiosis, producing four genetically distinct haploid cells, each with a single set of chromosomes (n). During the haploid phase, coccolithophores can reproduce both asexually and sexually. The fusion of two compatible haploid cells restores the diploid stage, completing their life cycle.
Mitosis. Mitosis is a process in which a single diploid parent cell (2n) divides once, producing two genetically identical daughter cells. This maintains the chromosome number, ensuring genetic continuity during growth, maintenance, and asexual reproduction.
Meiosis. Meiosis is a specialised form of cell division that produces haploid cells from a diploid parent cell and forms part of the sexual life cycle. During meiosis, a single diploid cell (2n) undergoes two successive divisions to produce four genetically distinct haploid cells (n), each containing a single set of chromosomes. By reducing the chromosome number and generating genetic variation through recombination and independent assortment, meiosis contributes to the genetic diversity of coccolithophore populations. The resulting haploid cells may continue to reproduce asexually by mitosis, giving rise to a haploid population. Eventually, some compatible haploid cells function as gametes and fuse through syngamy, restoring the diploid phase and completing the life cycle.
Syngamy (fusion of haploid gametes). In syngamy, two compatible haploid cells (n) fuse together, merging their cellular contents and nuclear material to create a diploid zygote. Afterwards, the zygote progresses into the diploid (2n) phase of the life cycle (Laguna et al., 2001). Evidence for syngamy mainly comes from ploidy analyses and observations of combination coccospheres, as direct observation of cell and nuclear fusion in culture conditions has yet to be achieved.
The non-motile diploid phase produces heterococcoliths composed of radially arranged calcite elements organised into complex structures. In contrast, the motile haploid phase produces holococcoliths, which consist of minute calcite crystallites, typically rhombohedra or hexagonal prisms, commonly less than 0.1 μm in size. The terms heterococcolith and holococcolith were introduced by Braarud et al. (1955).
During the motile haploid phase, coccolithophore cells possess two flagella and a haptonema, structures that are absent during the alternating non-motile diploid phase. Coccolith formation differs markedly between these life-cycle stages. Heterococcoliths are produced intracellularly within Golgi-derived coccolith vesicles, allowing precise biological control over crystal nucleation, growth, and morphology. They consist of relatively large calcite elements arranged in complex patterns, including laths, bars, wedges, and plates, which are assembled within predefined circular or elliptical frameworks to produce highly detailed structures (Varol, 2025a).
In the haploid phase, cells may be either naked or covered by holococcoliths. Holococcoliths were traditionally considered to form extracellularly with relatively limited biological control over crystal growth. However, Meyer and Taylor (2025) demonstrated that their formation may also occur intracellularly. Compared with heterococcoliths, holococcoliths possess simpler architectures composed of uniform calcite crystallites bound by an organic matrix. These crystallites are typically rhombohedral or hexagonal in form (Black, 1963; Gartner & Bukry, 1969; Kleijne, 1991), a microcrystalline structure most clearly observed using electron microscopy.
Under cross-polarised light (XPL), hexagonal prisms and rhombohedral crystals can be distinguished optically. In hexagonal prisms, the optical axis is parallel to the long axis of the crystal, whereas in rhombohedral crystals it is diagonal to the crystal faces (Varol, 2025a).
During transitions between life-cycle stages, coccolithophores may produce combination coccospheres bearing both heterococcoliths and holococcoliths. These forms provide important evidence linking the two phases of the life cycle. Identification and classification of holococcoliths, particularly in older sediments, remain challenging owing to their small size, relatively simple morphology, and susceptibility to diagenetic alteration.
In summary, coccolithophores alternate between a diploid phase that produces heterococcoliths and a haploid phase that may be naked or produce holococcoliths. The transition between these stages involves meiosis and syngamy, allowing both asexual reproduction and genetic recombination. This complex life cycle contributes to the ecological success and evolutionary diversity of coccolithophores throughout the world's oceans.
Carbon Cycling by Coccolithophores
Coccolithophores are photosynthetic microorganisms that inhabit the sunlit upper layers of the ocean. Through photosynthesis, they convert dissolved inorganic carbon (DIC), primarily dissolved carbon dioxide (CO₂), into organic biomass, known as particulate organic carbon (POC) (Passow & Carlson, 2012; Sigman & Hain, 2012). This process forms the basis of the biological carbon pump, through which a portion of the carbon fixed in surface waters is transported to deeper parts of the ocean and, in some cases, buried in marine sediments. As a result, coccolithophores contribute to the long-term seizure of carbon from the ocean-atmosphere system.
The Biological Carbon Pump
Through photosynthesis, coccolithophores convert dissolved inorganic carbon into organic biomass. When coccolithophore cells die, are consumed by zooplankton, or become incorporated into aggregates, their organic carbon begins a downward journey through the water column towards the seafloor (Heinze et al., 2015; Turner, 2015). Individual coccolithophore cells sink very slowly,, according to Bach et al. (2016), uncalcified Emiliania huxleyi cells sink at approximately 11 m year, whereas naked Calcidiscus spp. cells are estimated to sink at about 256 m year. At these rates, an uncalcified Emiliania huxleyi cell would require approximately 36 years to sink through a 400 m water column, whereas a naked Calcidiscus cell would require only about 1.6 years.
During their descent, many coccolithophore cells and particles are consumed by heterotrophic organisms or decomposed by bacteria, recycling much of their carbon back into seawater. Only a small fraction survives long enough to reach the seabed, where it may be consumed by benthic organisms (Turner, 2015) or become incorporated into marine sediments. Carbon that is buried within these sediments can remain isolated from the ocean-atmosphere system for thousands to millions of years, forming an important component of the long-term carbon cycle. Over time, deep-ocean circulation and upwelling return some dissolved carbon and nutrients to surface waters, where they can once again be used by photosynthetic organisms. This process, known as the biological carbon pump, plays an important role in regulating atmospheric carbon dioxide levels.
The Carbonate Pump
In addition to photosynthesis, coccolithophores produce coccoliths composed of calcium carbonate (CaCO₃). The formation of these coccoliths, a process known as calcification or biomineralization releases CO₂ within the cell, some of which may be reused during photosynthesis (Mackinder et al., 2012).
When coccolithophores die, their coccoliths sink through the water column. Some dissolve before reaching the seabed, releasing calcium ions and dissolved inorganic carbon back into seawater. This dissolved carbon may subsequently be transported by deep-ocean circulation and eventually returned to surface waters through upwelling. Other coccoliths reach the seafloor, where they accumulate and contribute significantly to calcareous marine sediments. The sinking rate of individual coccoliths is relatively slow. Isolated coccoliths of Calcidiscus leptoporus sink at approximately 0.74 m day (about 270 m year), implying that an individual coccolith could require about 1.5 years to sink through a 400 m water column (Bach et al., 2012). Comparable measurements for detached Emiliania huxleyi coccoliths are scarce, although their smaller size suggests substantially lower sinking velocities. Consequently, individual cells and coccoliths may require months to decades to traverse a 400 m water column, suggesting that efficient export to the seabed is likely to depend on aggregation into marine snow or incorporation into faecal pellets.
Coccolithophores are among the most important producers of pelagic carbonate sediments in the world's oceans (Hofmann et al., 2010; Renaud et al., 2002).
Over the course of geological period, buried carbonate sediments may be transformed into chalk, limestone, or marl through compaction and lithification (Folk, 1980). These rock formations can store carbon for millions of years before weathering, erosion, and tectonic processes eventually return it to the ocean-atmosphere system (Gibb, 1978).
Together, photosynthesis and calcification (biominaralisation) lift coccolithophores vital contributor in both the biological carbon pump and the marine carbonate cycle, linking the ocean, atmosphere, and geological record.
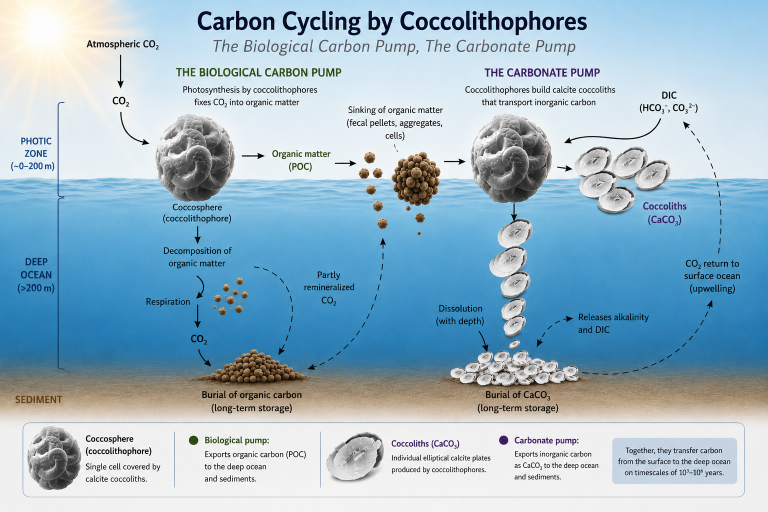
Simplified schematic presentation of carbon cycling by coccolithophores, illustrating the biological carbon pump and the carbonate pump, and their roles in the transfer of organic and inorganic carbon from the photic zone to the deep ocean and sediments.
References
Black, M. 1963. The fine structure of the mineral parts of coccolithophorids. Proceedings of the Royal Society of London. 174: 41-46.
Bach, L.T., Mackinder, L.C.M., Schulz, K.G., Wheeler, G., Schroeder, D.C., Brownlee, C. & Riebesell, U. 2016. Why marine phytoplankton calcify. Science Advances, 2(11): e1501822
Bach, L. T., Riebesell, U., Sett, S., Febiri, S., Rzepka, P., & Schulz, K. G. 2012. An approach for particle sinking velocity measurements in the 3–400 μm size range and considerations on the effect of temperature on sinking rates. Journal of Plankton Research, 34, 389–401.
Braarud, T., Deflandre, G., Halldal, P. & Kamptner, E. 1955. Terminology, nomenclature, and systematics of the Coccolithophoridae. Micropaleontology. 1(2): 157-159.
Falkowski, P. G. & Raven, J. A. 1997. Aquatic Photosynthesis. Princeton University Press.
Folk, R.L. 1980. Petrology of sedimentary rocks. Hemphill Publishing Company.
Gartner, S. & Bukry, D. 1969. Tertiary holococcoliths. Journal of Paleontology. 43(5): 1213-1221.
Gibb, J.G. 1978. Rates of coastal erosion and accretion in New Zealand. New Zealand Journal of Marine and Freshwater Research. 12(4): 429–456.
Heinze, C., Meyer, S., Goris, N., Anderson, L., Steinfeldt, R., Chang, N. … & Bakker, D.C. 2015. The ocean carbon sink–impacts, vulnerabilities and challenges. Earth System Dynamics. 6(1): 327–358.
Young, J. R., Geisen, M., Cros, L., Kleijne, A., Probert, I. & Ostergaard, J. B. 2003. A guide to extant coccolithophore taxonomy. Journal of Nannoplankton Research. S1: 1-132.