History of Calcareous Nannofossil Research
Early history of calcareous nannofossil Research, 1839 - 1900
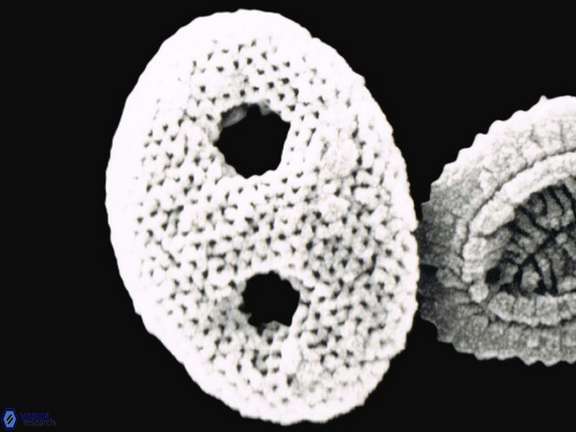
The scientific understanding of calcareous nannofossils developed gradually during the nineteenth century through a series of observations that transformed enigmatic microscopic particles into recognised components of living marine plankton. The foundations were laid by Ehrenberg (1839, 1843), who demonstrated that chalk deposits were largely composed of microscopic organisms, thereby establishing the concept that vast sedimentary rocks could originate from the accumulation of minute biological remains. Although coccoliths were not yet clearly distinguished, Ehrenberg’s work introduced the essential idea that microscopic life plays a major geological role.
A decisive step occurred when Huxley (1858), examining material recovered from deep-sea soundings in the North Atlantic, described minute calcareous plates and introduced the term coccolith. These structures were recognised as abundant constituents of marine sediments, but their biological nature remained uncertain. Around the same time, spherical bodies bearing coccoliths were observed and later termed coccospheres, although their relationship to living organisms was still debated.
The biological origin of coccoliths was first clearly proposed by Wallich (1861), who interpreted them as products of living marine organisms rather than inorganic precipitates. Later in the same year, Sorby (1861) confirmed their organic nature through microscopic investigation of chalk microstructures and acknowledged Wallich’s priority in recognising their biological origin. These studies marked the transition from viewing coccoliths as mineral curiosities to understanding them as biologically produced structures.
Subsequent investigations expanded morphological knowledge. Schmidt (1870) provided detailed descriptions of coccoliths and related forms, including elongate structures later termed rhabdoliths, contributing to early morphological classification. Wallich (1877) further clarified the structure of coccospheres, strengthening the interpretation that coccoliths formed external coverings of planktonic cells. During this period, distinct morphologies began to be recognised, including discoidal forms later known as discoliths and more complex shield-bearing types subsequently termed placoliths, although formal terminology developed gradually.
By the late nineteenth century, coccoliths were increasingly documented in modern marine environments. Observations by Joly and Dixon (1897) demonstrated their presence in coastal waters, indicating that coccolith-bearing organisms were active components of living plankton communities rather than exclusively deep-sea sedimentary remains. The decisive biological interpretation was achieved by Murray and Blackman (1898), who demonstrated that coccospheres and rhabdospheres represented living unicellular algae and that coccoliths formed skeletal elements produced by these organisms. This work effectively established the concept of the coccolithophore, a calcifying planktonic alga that produces coccoliths.
By the end of the nineteenth century, scientific understanding had progressed from the recognition of microscopic particles in chalk to the identification of coccolithophores as living planktonic algae whose coccoliths accumulated to form major marine sediments. This transformation laid the intellectual foundation for twentieth-century developments in nannoplankton research and calcareous nannofossil biostratigraphy.
The terminology of coccolith studies developed progressively during the nineteenth century. Huxley (1858) introduced the term coccolith for minute calcareous plates recovered from deep-sea sediments. Wallich (1861, 1877) subsequently recognised spherical aggregates of coccoliths, later termed coccospheres, and interpreted them as organised biological structures. Schmidt (1870) introduced the term rhabdolith for elongate forms, leading to recognition of morphologically distinct coccolith-bearing organisms such as Rhabdosphaera. As microscopy improved, discoidal and shield-bearing morphologies were distinguished, forming the basis of the later concepts of discoliths and placoliths. These terminological developments paralleled the transition from descriptive observation to biological and taxonomic understanding of coccolithophores.
Transition into 1900–1960
By 1900, coccoliths and coccospheres had moved from being enigmatic particles in deep-sea sediments to recognised biological structures associated with living plankton, culminating in the demonstration by Murray and Blackman (1898) that coccospheres represent living unicellular algae. The period 1900–1960 marks the transformation of this late-nineteenth-century insight into a coherent scientific discipline: coccolithophore biology and taxonomy were formalised, methodological advances (especially improved microscopy and later electron microscopy) revealed skeletal microstructure in unprecedented detail, and calcareous nannofossils became increasingly important in stratigraphy—particularly through the rising use of discoasters, nannoconids, and other key fossil groups in interregional correlation.
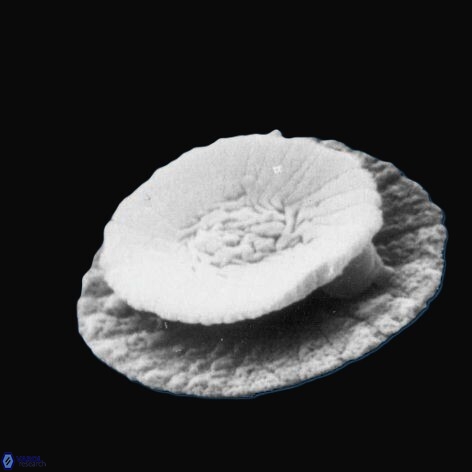
Summary of key developments in calcareous nannofossil research, 1900–1960

Early-twentieth-century research consolidated the biological interpretation of coccoliths and coccospheres and formalised the organisms that produce them. Work at the start of the century (e.g., Dixon, 1900; Ostenfeld, 1900; Blackman, 1902) continued to clarify coccosphere structure and coccolith formation, while Lohmann’s monograph (1902) was pivotal in establishing Coccolithophoridae as a coherent group of coccolith-producing flagellates and firmly embedding the term coccolithophore (coccolith-bearing organism) within plankton biology. Through Lohmann’s later methodological studies (1903, 1908, 1912, 1913, 1919), the concept of nannoplankton research was advanced by improved sampling and concentration methods, and coccolithophores became increasingly recognised as a routine component of marine phytoplankton investigations (e.g., Lemmermann, 1903, 1908; Steuer, 1903; Schiller, 1913–1916; Brunnthaler, 1911; Ostenfeld, 1910–1911; Murray & Hjort, 1912).
Between the 1920s and 1940s, a strong taxonomic and systematic framework developed alongside improvements in light microscopy and expanding geographic observations. Schiller’s syntheses (1925–1930) helped stabilise coccolithophore classification within algal systematics, while Kampfner’s extensive contributions from the late 1920s onward profoundly influenced both living and fossil coccolith taxonomy and morphology. His description of Nannoconus (Kamptner, 1931) recognised a distinctive rock-forming Mesozoic microfossil group and demonstrated the geological importance of calcareous nannofossils. A major terminological and taxonomic advance was the introduction of the genus Discoaster by Tan Sin Hok (1927), providing a formal framework for stellate nannoliths that later became among the most important markers in Cenozoic biostratigraphy. Concurrently, Deflandre’s studies during the 1930s–1950s expanded the fossil record and refined systematic concepts, including recognition of morphologically complex forms such as Schizosphaerella (Deflandre & Dangeard, 1938) and later Braarudosphaera (Deflandre, 1947). During this period, increasing recognition of discoidal coccoliths (later termed discoliths) and shield-bearing forms (placoliths) reflected improved understanding of skeletal architecture revealed by microscopy.
From the early 1950s onward, progress accelerated rapidly with the introduction of electron microscopy and efforts toward standardised terminology. Electron microscope investigations (e.g., Braarud, Gaarder, Markali & Nordli, 1952; Halldal, 1954; Halldal & Markali, 1954a-b, 1955; Deflandre & Fert, 1954; Downie & Honeycombe, 1956; Kamptner, 1950–1956) revealed coccolith microstructure at the crystallite scale, allowing taxonomy to be based increasingly on skeletal construction rather than external morphology alone. A landmark step toward conceptual and terminological standardisation was the synthesis by Braarud, Deflandre, Halldal and Kamptner (1955a-b), which clarified nomenclature and recognised two fundamentally different coccolith construction types, later termed heterococcoliths and holococcoliths, representing distinct biomineralisation modes that were subsequently interpreted as different phases within the coccolithophore life cycle and experimentally demonstrated by Parke and Adams (1960). Detailed structural analyses of fossil coccoliths (e.g., Black & Barnes, 1959-1975) further integrated biological and palaeontological perspectives.
Simultaneously, calcareous nannofossils emerged as powerful stratigraphic tools, particularly within petroleum geology. Bramlette and Riedel (1954) and Bramlette (1957) demonstrated the stratigraphic value of discoasters and related forms, while Stradner (1958–1960) developed discoaster-based correlations and recognised nannoplankton invasion events in sedimentary basins. By the end of the 1950s, systematic revisions and regional studies (e.g., Kamptner 1950-1964, Deflandre, 1950-1963; Brönnimann, 1955; Górka, 1957-1967; Noël, 1957–1990; Martini, 1958–1998; Stradner, 1958-2010; Manivit, 1959-1990; Vekshina, 1959; Trejo, 1959–1983; Brönnimann & Stradner, 1960) confirmed the global distribution and stratigraphic utility of calcareous nannofossils. Thus, between 1900 and 1960, coccolithophore biology, coccolith morphology and microstructure, and fossil-based stratigraphic application matured together, establishing the foundations for the rapid expansion of quantitative nannofossil biostratigraphy and palaeoceanographic research in the decades that followed.
Thus, by 1960, the conceptual, methodological, and taxonomic foundations of modern calcareous nannofossil research were firmly established, paving the way for the rapid development of high-resolution biostratigraphy and palaeoceanography in the following decades.
Establishment and Consolidation of Modern Calcareous Nannofossil Biostratigraphy, 1960–1980

Between 1960 and 1980, calcareous nannofossil research underwent its most profound transformation, evolving from a predominantly descriptive, taxonomic discipline into a mature, globally integrated biostratigraphic science. Advances in microscopy, taxonomy, stratigraphic methodology, ocean drilling, and biological research collectively established the conceptual and practical framework that continues to underpin modern nannofossil studies.
A fundamental technological turning point was the widespread adoption of transmission and scanning electron microscopy. Early investigations by Hay & Towe, 1962, 1963; Gaarder, 1962; Black, 1963; Reinhardt, 1964, Perch-Nielsen, 1967; Stradner & Adamiker, 1966; Stradner et al., 1968; Stradner & Edwards, 1968; Gartner (1968) and Bukry (1969) revealed the crystallographic organisation and constructional architecture of coccoliths with unprecedented clarity. These studies demonstrated that coccolith morphology reflected ordered crystal arrangement rather than simple calcite plates, allowing taxonomy to be redefined on structural principles rather than outline morphology alone. Hay & Sandberg, 1967 explicitly recognised the scanning electron microscope as a breakthrough for micropalaeontology, marking the beginning of modern morphological systematics.
At the same time, biological investigations clarified the formation of coccoliths and their life cycles. Research by Parke, Manton and co-workers, together with physiological experiments by Paasche (1962-1966), demonstrated biological control of calcification and documented alternation between coccolith-bearing and non-calcifying phases in coccolithophores. These discoveries firmly linked fossil coccoliths to the biology of living phytoplankton and to oceanographic processes.
The early 1960s also marked the emergence of systematic nannofossil biostratigraphy. The pioneering studies of Bramlette & Sullivan 1961 and Sullivan 1964 demonstrated the stratigraphic value of nannoplankton assemblages in Early Tertiary successions. Martini & Bramlette 1964 recognised major evolutionary turnover across the Maastrichtian–Danian boundary, illustrating the sensitivity of nannofossils to global environmental change. Parallel investigations by Stradner (1962, 1963) documented the stratigraphic distribution of discoasters and other key taxa. At the same time, Bramlette & Wilcoxon 1967, Hay et al. (1966), Hay et al. (1967), Hay and Mohler (1967), and Gartner (1969) refined Tertiary taxonomy and stabilised important Discoaster species concepts that became fundamental to later zonations.
During the mid-1960s, the rapid expansion of regional studies extended geographic and stratigraphic coverage worldwide. Reinhardt (1964-1972) established robust systematic frameworks for Mesozoic and early Cenozoic taxa, while Noël (1959- 1965) provided a comprehensive classification of Jurassic coccoliths that became a cornerstone of later taxonomy. Contributions by Gartner (1968), Roth (1968), Perch-Nielsen (1968-1969), Martini (1968-1969), Bukry & Bramlette (1968-1969) further refined stratigraphic applications across the world. Electron-microscope studies by Perch-Nielsen during the late 1960s introduced ultrastructural criteria that significantly improved higher-level classification and taxonomic consistency.
A decisive conceptual advance followed with the development of formal biostratigraphic zonations. The synthesis by Hay, Mohler, Roth, Schmidt & Boudreaux 1967 established a coherent Cenozoic nannoplankton zonation for the Gulf Coast and Caribbean region and demonstrated reliable transoceanic correlation. Roth (1968, 1970) expanded these concepts into globally applicable zonal frameworks, while Martini 1971 formalised standard Palaeogene and Neogene calcareous nannoplankton zonations that rapidly became international reference standards. Parallel work by Bukry (1971-1975) introduced alternative zonal schemes that were widely adopted in industrial stratigraphy, particularly in petroleum exploration (Okada & Bukry, 1980).
The late 1960s and early 1970s coincided with the emergence of deep-sea drilling programmes, which transformed the scale of nannofossil research. Studies associated with early ocean drilling (e.g., Bukry & Bramlette 1969; Hay 1970; Bukry 1971) demonstrated that nannofossil successions were globally synchronous and capable of providing high-resolution age control in pelagic sediments. These results confirmed calcareous nannofossils as one of the most powerful tools for marine stratigraphy and enabled precise correlation across ocean basins.
In 1969, Maria Farinacci began producing the Nannofossil Catalogue, an extraordinarily ambitious and demanding undertaking. Over the following two decades, she compiled 13 volumes (1969–1989), systematically documenting the taxonomy and literature of calcareous nannofossils. The catalogue rapidly became an indispensable reference work and effectively served as the primary handbook for nannofossil taxonomists worldwide.
Simultaneously, evolutionary and phylogenetic perspectives began to influence taxonomy. Gartner (1970) and Romein (1979) emphasised lineage concepts and evolutionary succession, linking taxonomy directly with stratigraphic interpretation. Haq (1966-1968) contributed significantly to quantitative and stratigraphic analyses of Paleogene assemblages, while Okada & Honjo (1973a-b, 1975), Okada & McIntyre (1977), Okada & Thierstein (1979) and Thierstein (1976, 1977) later advanced quantitative ecological and palaeoceanographic approaches, integrating assemblage analysis with oceanographic interpretation. Romein (1979) refined biostratigraphic calibration across key Paleogene intervals, strengthening interregional correlation.
Throughout the 1970s, systematic refinement and taxonomic stabilisation continued. Perch-Nielsen (1971a-1979) produced influential syntheses that integrated morphology, taxonomy, and stratigraphy across the Mesozoic and Cenozoic, while Bukry (1971-1975), Gartner (1967-1974), and Müller (1970-1977) contributed substantially to nannofossil zonation and regional calibration. The continued work of Stover (1966), Worsley (1971), Black (1971-1972), Wise & Wind (1977), Sissingh (1977) and Roth (1978) further strengthened Mesozoic taxonomy and classification schemes.
Another important milestone occurred in 1977, when the International Nannoplankton Association (INA) was founded under the leadership of Dr Katharina Perch-Nielsen. In the same year, the INA Newsletter was established to facilitate communication among nannopalaeontologists and to provide a rapid means of disseminating research results. The newsletter later evolved into the Journal of Nannoplankton Research, which continues to serve as the principal publication venue for the international nannoplankton community.
Equally important was the organisation and accessibility of the rapidly expanding literature. The annotated bibliographies of Loeblich & Tappan (1963–1970) provided an essential global index of calcareous nannoplankton research, serving as the primary reference framework for several decades. Subsequent indexing efforts by Van Heck further systematised nannofossil taxonomy and literature, greatly facilitating comparative and applied studies.
By the late 1970s, calcareous nannofossil biostratigraphy had reached full disciplinary maturity. Electron microscopy had stabilised morphological taxonomy; biological studies clarified coccolith formation; global zonation schemes enabled high-resolution correlation; and ocean drilling confirmed worldwide stratigraphic synchrony. The discipline had evolved into a quantitative and predictive stratigraphic tool widely applied in academic research and hydrocarbon exploration.
This period also established the intellectual environment in which a new generation of biostratigraphers further refined applied nannofossil biostratigraphy, integrating taxonomy, evolutionary concepts and stratigraphic interpretation into operational geological practice. By 1980, the essential methodological and conceptual foundations of modern calcareous nannofossil research were firmly in place, enabling the subsequent development of integrated stratigraphy and palaeoceanographic reconstruction in the following decades.
Publication Trends and Milestones in Calcareous Nannofossil Research
